Il citoscheletro consiste in una rete mutevole dinamicamente di tre diversi filamenti proteici nel citoplasma delle cellule.
Danno alla cellula e alle strutture intracellulari organizzative come gli organelli e le vescicole struttura, stabilità e mobilità intrinseca (motilità). Alcuni dei filamenti sporgono dalla cellula per supportare la motilità della cellula o il trasporto diretto di corpi estranei sotto forma di ciglia o flagelli.
Cos'è il citoscheletro?
Il citoscheletro delle cellule umane è costituito da tre diverse classi di filamenti proteici. I microfilamenti (filamenti di actina) con un diametro da 7 a 8 nanometri, costituiti principalmente da proteine di actina, servono a stabilizzare la forma cellulare esterna e la motilità della cellula nel suo complesso, nonché le strutture intracellulari.
Nelle cellule muscolari, i filamenti di actina consentono ai muscoli di contrarsi in modo coordinato. I filamenti intermedi, che hanno uno spessore di circa 10 nanometri, forniscono anche resistenza meccanica e struttura per la cellula. Non sono coinvolti nella motilità cellulare. I filamenti intermedi sono costituiti da varie proteine e dimeri delle proteine, che si combinano per formare fasci avvolti come corde (tonofibrille) e sono strutture estremamente resistenti allo strappo. I filamenti intermedi possono essere suddivisi in almeno 6 tipi diversi con compiti diversi.
La terza classe di filamenti è costituita da minuscoli tubi, i microtubuli, con un diametro esterno di 25 nanometri. Sono costituiti da polimeri di dimeri di tubulina e sono i principali responsabili di tutti i tipi di motilità intracellulare e della motilità delle cellule stesse. Per supportare la mobilità delle cellule, i microtubuli sotto forma di ciglia o flagelli possono formare processi cellulari che sporgono dalla cellula. La rete dei microtubuli è per lo più organizzata dal centromero ed è soggetta a cambiamenti estremamente dinamici.
Anatomia e struttura
I gruppi di sostanze microfilamenti, filamenti intermedi (IF) e microtubuli (MT), tutti e tre assegnati al citoscheletro, sono quasi onnipresenti all'interno del citoplasma e anche all'interno del nucleo cellulare.
Gli elementi costitutivi di base dei filamenti di micro o actina negli esseri umani sono costituiti da 6 proteine isoforme di actina, ciascuna delle quali differisce solo per pochi amminoacidi. La proteina actina monomerica (G-actina) lega il nucleotide ATP e forma lunghe catene molecolari di monomeri di actina, ciascuno dei quali separa un gruppo fosfato, di cui due si connettono per formare filamenti di actina elicoidali. I filamenti di actina nei muscoli lisci e striati, nei muscoli cardiaci e nei filamenti di actina non muscolari differiscono leggermente l'uno dall'altro. L'accumulo e la scomposizione dei filamenti di actina sono soggetti a processi molto dinamici e si adattano ai requisiti.
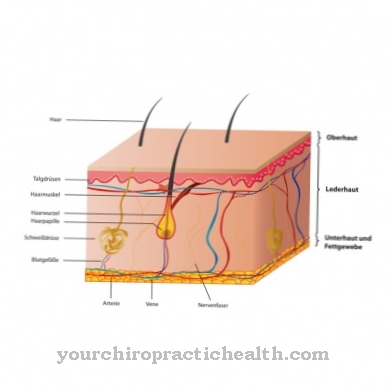
I filamenti intermedi sono costituiti da varie proteine strutturali e hanno un'elevata resistenza alla trazione con una sezione trasversale di circa 8-11 nanometri. I filamenti intermedi sono suddivisi in cinque classi: cheratine acide, cheratine basiche, tipo desmina, neurofilamenti e tipo lamina. Mentre le cheratine si trovano nelle cellule epiteliali, i filamenti di tipo desmina si trovano nelle cellule muscolari dei muscoli lisci e striati, nonché nelle cellule del muscolo cardiaco. I neurofilamenti che sono presenti praticamente in tutte le cellule nervose sono composti da proteine come Internexina, Nestina, NF-L, NF-M e altre. Filamenti intermedi del tipo laminare si trovano in tutti i nuclei cellulari all'interno della membrana nucleare nel carioplasma.
Funzione e compiti
La funzione e i compiti del citoscheletro non sono affatto limitati alla forma strutturale e alla stabilità delle cellule. I microfilamenti, che si trovano principalmente in strutture a rete direttamente sulla membrana plasmatica, stabilizzano la forma esterna delle cellule. Ma formano anche protuberanze di membrana come gli pseudopodi. Le proteine motorie, da cui vengono costruiti i microfilamenti nelle cellule muscolari, assicurano le contrazioni necessarie dei muscoli.
I filamenti intermedi ad altissima resistenza alla trazione sono di massima importanza per la resistenza meccanica delle celle. Hanno anche una serie di altre funzioni. I filamenti di cheratina delle cellule epiteliali sono collegati tra loro meccanicamente indirettamente tramite desmosomi, in modo che il tessuto cutaneo riceva una forza bidimensionale simile a una matrice. Gli IF sono collegati agli altri gruppi di sostanze nel citoscheletro tramite proteine associate ai filamenti intermedi (IFAP), assicurano un certo scambio di informazioni e la resistenza meccanica del tessuto corrispondente. Questo crea strutture ordinate all'interno del citoscheletro. Enzimi come chinasi e fosfatasi assicurano che le reti siano costruite, ristrutturate e scomposte rapidamente.
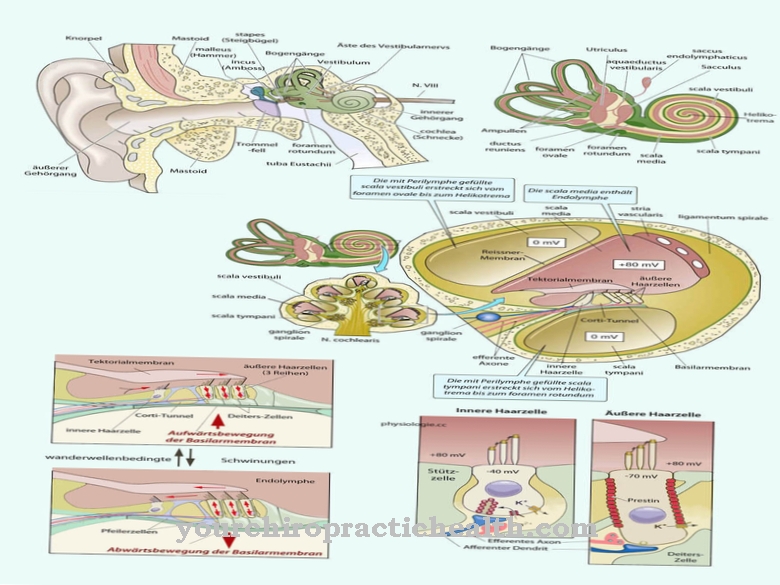
Diversi tipi di neurofilamenti stabilizzano il tessuto nervoso. Le lamine controllano la rottura della membrana cellulare durante la divisione cellulare e la sua successiva ricostruzione. I microtubuli sono responsabili di compiti come il controllo del trasporto di organelli e vescicole all'interno della cellula e l'organizzazione dei cromosomi durante la mitosi. Nelle cellule in cui i microtubuli formano microvilli, ciglia, flagelli o flagelli, i MT assicurano anche la motilità dell'intera cellula o si occupano della rimozione di muco o corpi estranei tali. B. nella trachea e nel condotto uditivo esterno.
Puoi trovare i tuoi farmaci qui
➔ Medicinali contro i disturbi della memoria e l'oblioMalattie
Disturbi nel metabolismo del citoscheletro possono derivare da difetti genetici o da tossine fornite esternamente. Una delle malattie ereditarie più comuni associate a un'interruzione nella sintesi di una proteina di membrana per i muscoli è la distrofia muscolare di Duchenne.
Un difetto genetico impedisce la formazione della distrofina, una proteina strutturale necessaria nelle fibre muscolari dei muscoli scheletrici striati. La malattia si manifesta nella prima infanzia con un decorso progressivo. Anche le cheratine mutate possono avere effetti gravi. L'ittiosi, la cosiddetta malattia del pelo di pesce, porta all'ipercheratosi, uno squilibrio tra la produzione e l'esfoliazione delle scaglie cutanee, dovuto a uno o più difetti genetici sul cromosoma 12. L'ittiosi è la più comune malattia ereditaria della pelle e richiede una terapia intensiva, che però può solo alleviare i sintomi.
Altri difetti genetici che portano a un'interruzione del metabolismo dei neurofilamenti causano z. B. sclerosi laterale amiotrofica (SLA). Alcune micotossine note (tossine fungine) come quelle della muffa e degli agarichi volanti interrompono il metabolismo dei filamenti di actina. La colchicina, la tossina del croco autunnale, e il taxolo, che si ottiene dagli alberi di tasso, sono usati specificamente per la terapia del tumore. Intervengono nel metabolismo dei microtubuli.









.jpg)

.jpg)
.jpg)